


氮污染是环境领域的一个严重问题,污水处理厂(wastewater treatment plants,WWTPs)广泛采用活性污泥法进行脱氮。活性污泥系统由复杂微生物群体构成,其中细菌群落在保持污水处理的稳定性方面发挥着重要作用[1],脱氮功能微生物如氨氧化细菌(ammonia-oxidizing bacteria,AOB)、亚硝酸盐氧化细菌(nitrite-oxidizing bacteria,NOB)和反硝化细菌(de⁃ nitrifying bacteria,DNB)等在脱氮过程中发挥重要作用,另外,脱氮相关功能基因与脱氮性能之间也存在显著相关性[2]。微生物群落依赖于不同分类群之间的相互作用,其中枢纽属或关键属对群落的稳定性很重要[3],有研究人员提出微生物全尺度分类法[4],可将微生物分为6类:丰富菌属(abundant taxa,AT)、中丰度菌属(moderatetaxa,MT)、稀有菌属(rare taxa,RT)、条件丰富菌属(conditionally abundant taxa,CAT)、条件丰富或稀有菌属(conditionally rare or abundant taxa,CRAT)和条件稀有菌属(conditionally rare taxa,CRT),而目前研究多聚焦于相对丰度较高的菌属,而忽略了依然有重要作用的稀有类群[4]。实际污水成分复杂,工业生产中出现的多种新污染物会在WWTPs中富集并对活性污泥体系产生影响,同时这些新污染物会形成复合污染,发生不同的相互作用对活性污泥产生更为复杂的影响。如抗生素与重金属复合污染会对活性污泥体系硝化性能、微生物群落结构、脱氮功能微生物和功能基因等产生影响,并且研究发现CRAT是微生物群落中的核心分类单元[5]。因此,探究新污染物复合污染胁迫对活性污泥系统中微生物群落的影响具有重要意义。
卤代咔唑(polyhalogenated carbazoles,PHCs)是一类具有潜在持久性、生物累积性和毒性的物质[6],目前已在国内外多数水域中检测到[7-9]。三氯咔唑(3-chlorocarbazole,3-CCZ)是其中最常见的PHCs种类,具有类二噁英毒性,试验发现其会导致斑马鱼体内一些基因的表达发生变化并且激活芳香烃受体(AhR)致使心脏畸形[10-11],但这些研究集中于模式生物,关于PHCs对活性污泥的影响尚不清楚。纳米材料(nanoparticles,NPs)特别是铁基NPs由于其独特的性能被大量利用,Fe3O4 nanoparticles(Fe3O4 NPs)是其中受到关注较多的材料并且被发现对废水处理过程有重大的影响[12-13]。Ma等[14]发现Fe3O4 NPs可以显著改变活性污泥中亚硝化单胞菌(Nitrosomonas)、亚硝化螺旋菌(Nitrosospira)和硝化螺旋菌(Nitrospira)等功能菌的相对丰度从而影响活性污泥体系中氨氧化和亚硝酸盐氧化过程;Xu等[15]发现Fe3O4 NPs可以提高厌氧氨氧化相关功能菌的活性。然而这些研究多聚焦于Fe3O4 NPs单一污染,它与其他污染物的复合污染对活性污泥系统的影响研究则较少。因此,有必要探讨PHCs和Fe3O4 NPs复合污染对活性污泥系统微生物群落的影响。
基于上述论点,本文研究了不同浓度3-CCZ与Fe3O4 NPs形成的复合污染对活性污泥体系中细菌群落的影响,并且探究了低温环境下复合污染胁迫对体系的影响。主要研究内容如下:(1)基于全尺度分类探索活性污泥微生物群落的响应;(2)全尺度分类下6个类群间的相互作用;(3)揭示复合污染胁迫对脱氮功能属和功能基因的影响。本研究为探索3-CCZ与Fe3O4 NPs复合污染提供了新视角,以期为推动PHCs与NPs复合污染的环境风险评价提供理论基础。
1 材料与方法
1.1 活性污泥微宇宙的构建与试验设计
本研究分为2个阶段,3-CCZ的环境浓度为ng/L级别[6, 16-17],考虑到其累积性[18],选择了2种浓度:低浓度(L:0.05 mg/L)和高浓度(H:5 mg/L)。第一阶段是考察3-CCZ和Fe3O4 NPs复合污染对活性污泥短期胁迫的影响,投加1 mg/L Fe3O4 NPs和不同浓度的3- CCZ(0.05和5 mg/L)到活性污泥反应器中,在指定操作温度(22±1)℃下运行12个周期。第二阶段为考察低温环境下3-CCZ与Fe3O4 NPs复合污染胁迫对活性污泥系统微生物的影响,各体系中污染物添加浓度不变,操作温度改为(12±1)℃并运行13个周期。每个周期以体系内NH4+-N降解98%以上为标志,停止曝气。不含任何污染物的体系和只含Fe3O4 NPs的体系在胁迫阶段命名为Control和NPs,同时含有3-CCZ和Fe3O4 NPs的体系根据3-CCZ浓度依次被命名为CCZ_L和CCZ_H,这些体系在低温阶段对应名称分别为Control_2、NPs_2、CCZ_L_2和CCZ_H_2。3-CCZ和Fe3O4 NPs均购自上海麦克林生化有限公司。
以500 mL锥形瓶为反应器,工作体积为250 mL,包括125 mL活性污泥和125 mL人工配水,活性污泥取自北京市某城市污水处理厂曝气池,颜色呈黄褐色,污泥浓度(mixed liquor suspended solids,MLSS)控制在5000 mg/L,污泥沉降比SV30为39%,SVI为59 mL/g。人工配水组成如下:葡萄糖0.562 g/L,氯化铵0.268 g/L,碳酸氢钠0.840 g/L,磷酸二氢钾0.030 g/L,氯化钠0.060 g/L,氯化钾0.010 g/L,微量元素1 mL/L。通过曝气体系供氧以保证体系的溶解氧(DO)保持在2 mg/L以上,每个体系的pH值保持在7.5~8.5,通过水浴恒温振荡器使每个体系的温度保持在每个阶段需要的温度范围,通过多参数水质仪(WTW Multi3630 IDS)对体系中的操作条件进行连续监测。
1.2 DNA提取和测序分析
在第14周期和第27周期结束时从反应器中收集污泥样品,共得到8个样品:Control、NPs、CCZ_L、CCZ_H、Control_2、NPs_2、CCZ_L_2和CCZ_H_2。利用E.Z.N.A. ®Soil DNA试剂盒(Omega Bio-tek,USA)提取基因组DNA,并用NanoDrop-2000(Thermo Fish⁃ er Scientific,USA)测定DNA浓度。使用lllumina MiSeq PE300测序仪(lllumina,USA)进行细菌16SrRNA基因(V3~V4区)测序(美吉生物云平台)。将有效序列按照相似度>97% 划分到相同的操作分类单元(operational taxonomic units,OTUs)。通过RDP数据库对OTUs进行物种注释。原始数据已上传至国家生物技术信息中心(NCBI)基因数据库(PRJNA1059288)。利用Shannon指数和Chao1指数评估微生物多样性和丰富度,同时,采用主成分分析(principal component analysis,PCA)探究微生物群落结构。
1.3 全尺度分类与功能属
采取全尺度分类方法[4]将细菌划分为6个类群,其中,菌属在所有样品中的相对丰度均在1%以上定义为AT,菌属在部分样品中相对丰度在1% 以上,但均不低于0.1% 定义为CAT;菌属在所有样品中的相对丰度在0.1%~1%之间定义为MT;菌属在所有样品中相对丰度均低于1%,在部分样品中低于0.1%定义为CRT;菌属在所有样品中相对丰度均低于0.1% 定义为RT;菌属在样品的相对丰度从低于0.1%到高于1%都存在定义为CRAT。
根据先前的研究[19],选择了脱氮功能属,包括氨氧化细菌(AOB)、亚硝酸盐氧化细菌(NOB)和反硝化细菌(DNB)进行详细分析。通过搜索ko00910途径中的特定京都基因和基因组同源百科全书(KOs)分析了参与硝化和反硝化的功能基因,并应用热图来研究功能基因的出现以及变化情况。
1.4 网络分析
建立相关性网络以研究微生物菌属在不同条件下的相互作用。使用R(V4.3.1)计算了微生物菌属之间的Spearman等级相关系数(SRCCs),构建相关矩阵,P≤0.05定义为显著相关,使用Gephi(V0.92)展示微生物菌属(6大类)之间的网络关系。
2 结果与分析
2.1 微生物群落多样性与结构
各样品的多样性指数和丰富度指数变化如图 1(a)所示,无论是胁迫阶段还是低温阶段,各样品间Shannon指数的变化范围在10%以内,说明各样品中微生物群落多样性变化较小,污染物的添加对活性污泥体系微生物群落多样性的影响不显著。对于Chao1指数,添加了污染物的体系中Chao1指数下降,复合污染更为明显尤其是CCZ_L,这说明污染物的添加降低了微生物群落的丰富度,3-CCZ与Fe3O4 NPs复合污染对微生物可能存在协同作用。在低温阶段,NPs_2的Chao1指数从1060下降至940,这可能是因为Fe3O4 NPs在活性污泥体系中会溶解出离子而对微生物产生毒性[20],在复合污染体系中Chao1指数则上升,进一步证明了3-CCZ与Fe3O4 NPs之间存在相互作用,在低温环境下呈现出拮抗作用。总之,污染物的添加对活性污泥体系中微生物群落丰富度会有不同程度的影响,但不会影响微生物多样性,并且3-CCZ与Fe3O4 NPs的复合污染在常温与低温下对体系微生物丰富度产生了不同的作用。
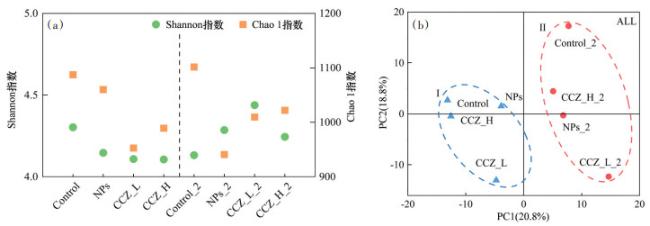
通过PCA对复合污染下微生物的群落结构进行分析,如图 1(b)所示,整个微生物群落可分为2个组别: Ⅰ(Control、NPs、CCZ_L和CCZ_H)和Ⅱ(Control_ 2、NPs_2、CCZ_L_2和CCZ_H_2),同一个阶段的样品聚在一起,说明了低温对微生物群落结构的改变。在Ⅰ组中,CCZ_L距离Control最远,NPs与CCZ_H则与Control距离较近,可能是Fe3O4 NPs溶解产生的金属离子会促进细菌分泌胞外聚合物[21],对自身形成保护导致单一Fe3O4 NPs污染对微生物群落结构的影响较小。在Ⅱ组中,添加了污染物的体系与Contorl距离均较远,其中CCZ_L_2距离最远,相较于胁迫阶段,低温阶段中添加了污染物的体系微生物群落结构与Control差别更大。
2.2 丰富属和稀有属的响应
基于微生物全尺度分类,共得到6类细菌菌群(AT、CAT、CRT、CART、MT和RT)。AT(39.04%~ 53.21%)与CAT(32.26%~45.90%)是整个微生物群落中占主导地位的微生物类群,其次是CRAT(7.34%~ 10.74%)和CRT(2.97%~3.72%),MT(1.83%~3.17%)与RT(0.50%~0.76%)占比极少;NPs、CCZ_L和CCZ_H中AT相对丰度高于Control,污染物的添加促进了体系中AT的生长,在低温阶段仅有CCZ_L_2中AT相对丰度下降,说明低浓度3-CCZ与Fe3O4 NPs复合污染在低温下对AT产生了抑制作用;对于CAT,添加了污染物的体系中相对丰度均下降,说明CAT对污染物较敏感,低温阶段各体系相对丰度均有所下降,其中CCZ_L_2的变化幅度最小(从35.76% 下降至35.15%),可能是低温对体系中CAT产生了抑制作用,而低浓度3-CCZ与Fe3O4 NPs复合污染的胁迫则对该作用有所减缓。
为进一步研究,把菌群分为丰富类和稀有类2大类。如 图 2(b)所示Saccharimonadales(39.04%~ 53.21%)是AT中的唯一菌属,它是一种反硝化细菌[22],可在部分反硝化中发挥关键作用[23]。胁迫阶段试验组Saccharimonadales相对丰度均高于对照组,可能是污染物会对Saccharimonadales的生长起到促进作用,这也表明了经过污染物的胁迫可能会促进活性污泥系统的反硝化能力的改善,而CCZ_L和CCZ_H中相对丰度低于NPs,可能是3-CCZ与Fe3O4 NPs存在拮抗作用。经过低温阶段Control_2中该菌属的相对丰度从34.53%上升至46.57%,而剩余体系中它的相对丰度则下降,可能是低温更适合该菌的生长,而3-CCZ与Fe3O4 NPs在低温下则对其产生了抑制。
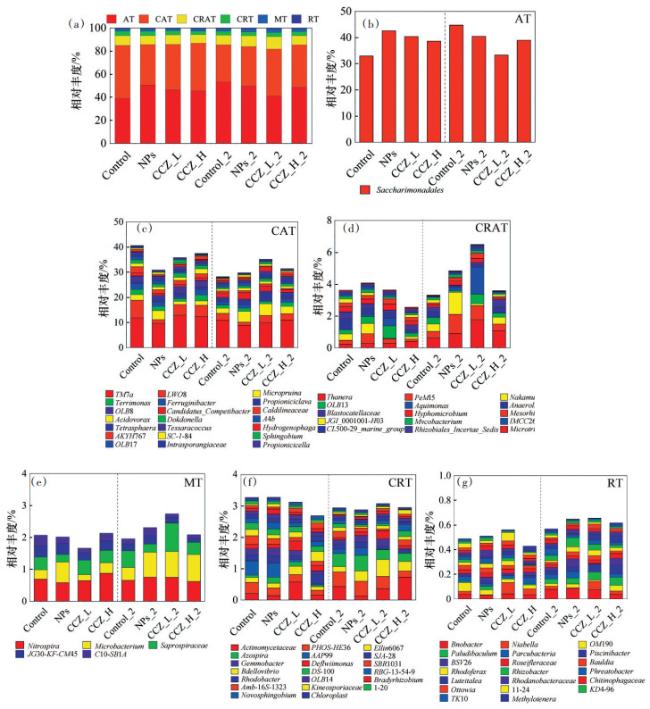
在CAT中(图 2(c))胁迫阶段各体系微生物相对丰度整体高于低温阶段,而在不同阶段内试验组与对照组微生物相对丰度的趋势则相反。胁迫阶段Control内微生物相对丰度高于其他体系,在低温阶段Control_2中CAT的相对丰度则从40.60% 下降至28.24% 并且低于试验组,这表明CAT更易受低温影响。TM7a(8.90%~12.86%)是CAT中绝对的优势属,其已经在人类样本与活性污泥中发现[24],它在各样本中的相对丰度变化较小,可能是其对3-CCZ与Fe3O4 NPs不敏感。有部分菌属如OLB8与CAT整体的变化趋势相似,在低温阶段相对丰度减少,OLB8参与了污水处理厂中的不同碳化合物的降解和磷与氮的去除[25]。另外一部分菌属如Micropruina和Candidatus_Competibacter等则在低温阶段相对丰度上升,某些添加了污染物的体系菌属的相对丰度上升幅度高于Control_2。
在CRAT中(图 2(d)),没有一种菌属一直占主导地位。在胁迫阶段,CCZ_H中CRAT的相对丰度是最低的,CCZ_L和NPs中CRAT相对丰度则与Con⁃ trol相近,但各样本中的主体菌属不同,在NPs中占比最多的是PeM15(0.61%)和硝化螺旋菌属(Nakamurella,0.65%),CCZ_L中则是OLB13(0.44%)和水单胞菌属(Aquimonas,0.12%),这说明CRAT对污染物比较敏感导致各样本中占主导地位的菌属不同。经过低温阶段后试验组中CRAT相对丰度均有所上升,陶厄氏菌属(Thauera)与Pem15是各体系中均有所增长的菌属,Thauera在自养反硝化和异养反硝化过程中都具有重要作用并且其丰度会受Fe3O4 NPs浓度的影响[22, 26-27]。除此之外各体系中相对丰度变化最大的菌属有所不同,例如NPs_2中的Nakamurella(0.65% 上升至1.39%)和CCL_L_2中的Aquimonas(0.12% 上升至0.21%),说明低温下3-CCZ与Fe3O4 NPs会促进CRAT的生长,并且不同浓度3-CCZ与Fe3O4 NPs的复合污染对菌属的影响不同。
MT(图 2(e))中含5个属:Nitrospira(0.59%~ 0.88%)、硝化螺旋菌属(Microbacterium,0.19%~ 0.85%)、腐败螺旋菌属(Saprospiraceae,0.23%~ 0.89%)、JG30-KF-CM45(0.13%~0.35%)和C10- SB1A(0.11%~0.33%),占主导地位的是Nitrospira,它是硝化微生物中种类最多的一种,在自然和工程环境的硝化作用中具有重要意义[27-28],其相对丰度的变化也较小,说明Nitrospira对3-CCZ与Fe3O4 NPs并不敏感。Microbacterium与Saprospiraceae是各样本中变化较大的菌属,不同样本中其变化情况各不相同,相似的是低温下污染物的添加可以促进其生长,尤其是低浓度3-CCZ与Fe3O4 NPs的复合污染。JG30- KF-CM45与C10-SB1A的变化则较小。
在本研究中,有98个属被分类为CRT,选择前20的属进行分析(图 2(f))。没有一种菌属一直占有主导地位,且各菌属在不同样品中的分布规律并不明显,这说明一部分稀有菌属对污染物比较敏感,受到影响比较明显,如CCZ_L与CCZ_H中SBR1031、AAP99和PHOS-HE36等菌属相对丰度小于Control,而蛭弧菌属(Bdellovibrio)和Defluviimonas等菌属则高于对照,另外还有一些菌属会在某个体系中成为优势菌属,如CCZ_L中的放线菌属(Actinomycetaceae)。
对于RT(图 2(g)),分析了503个属中的前20个,每个属在不同样本中的分布是不规则的,进一步说明了稀有属对污染物更为敏感,与CRT不同的是,低温下污染物的添加使得RT类相对丰度有所上升,如Paludibaculum、红育菌属(Rhodoferax)和Piscinibacter等菌属在NPs_2、CCZ_L_2和CCZ_H_2中都高于其胁迫阶段;另外还有一些菌属在某个体系内相对丰度会有增长,如CCZ_L_2中的Niabella和CCZ_H_2中的BSV26等。总的来说,稀有属微生物对污染物的添加更为敏感,而丰富属微生物则对不同浓度的复合污染更为敏感,可能是污染物间存在的不同种相互作用,另外复合污染的胁迫也会改变某些菌属对低温环境的响应。
2.3 丰富属和稀有属间交互作用
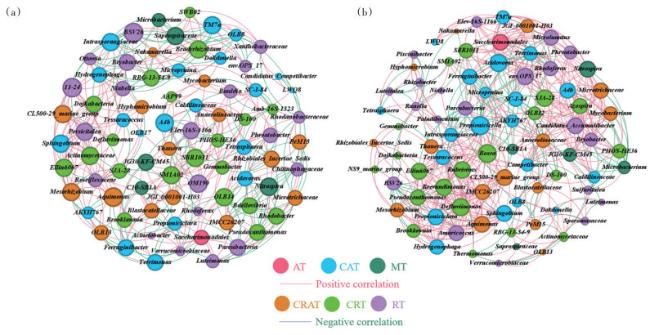
自然生态系统中的微生物并不是作为个体种群单独存在的。相反,它们相互作用,形成复杂的微生物群落[29]。本研究通过网络分析探究微生物间交互作用(图 3)。胁迫阶段和低温阶段的交互网络中分别有391和502条共现关系,正相关占比分别为61.64% 和76.49%,表明微生物之间多为共现模式并且低温下微生物之间的交互作用更为复杂。胁迫阶段中CRT与其他类群有最多的共现关系(160条),其中CRT与RT的相关关系最为明显(52条),正相关关系达到57.69%,这说明CRT与RT有着重要地位。稀有属在6个分类群中也很重要,在之前的研究中Zhao等[5]发现抗生素与重金属复合污染胁迫下CRT和RT与其他类群也存在复杂的网络关系。低温阶段CAT与其他类群有最多的相关关系(148条),表明CAT是处于微生物交互网络关键生态位的类群。虽然在2个阶段中AT与RT之间的共现关系不多(< 6条),但CAT与RT、CRAT及CRT之间的关系线数量较大(共305条),其中61.90% 为正相关,表明丰富属与稀有属之间存在着复杂的关联。此外,还对6个细菌分类群的核心属进行了探索。在胁迫阶段中,网络关系前10的属中各类群分布如下:3个CRT、3个CRAT、2个CAT和2个RT,其中CRT与其他类群的共现关系最多,Actinomycetaceae、Bradyrhizobium和Ellin6067是CRT的中心属。在低温阶段,网络关系前10的属中有5个属于CRT,博斯氏菌(Bosea)是CRT的中心属,拥有最多的相关关系。Aquimonas(属于CRAT)在2个阶段中都是网络关系前10的属。总之,CAT和CRT是核心分类单元,与其他分类单元有着复杂的相互作用,同时,稀有类群与其他类群也存在复杂的网络关系。
2.4 功能微生物和功能基因
对2个阶段中的脱氮功能属进行讨论。本研究中检测到的AOB是Nitrosomonas(0.00%~0.10%)(图 4(a)),Control中未检测出该菌属,其余体系中Nitrosomonas相对丰度有所增长尤其是CCZ_H,可能复合污染物可以促进其生长并且Nitrosomonas对高浓度3-CCZ与Fe3O4 NPs的复合污染更敏感。Ma等[14]发现Fe3O4 NPs可以显著引起活性污泥体系中Nitrosomonas相对丰度的变化,Nitrospira与氨氧化成亚硝酸盐的过程有关[30],复合污染可以通过影响其相对丰度从而影响氨氧化过程。低温阶段情况相似,CCZ_H_2中该菌属相对丰度从0.1%降低到0.03%,变化最大。Nitrospira(0.58%~0.88%)属于NOB,其与亚硝酸盐氧化成硝酸盐的过程密切相关[31],2个阶段中各样本NOB的相对丰度变化都较小(图 4(b)),说明Nitrospira对3-CCZ与Fe3O4 NPs在短期内并不敏感。
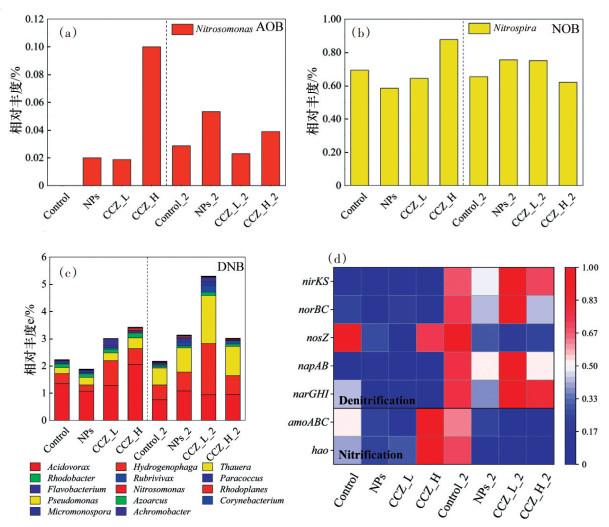
DNB是占比最高的脱氮功能微生物(1.88%~ 7.04%)(图 4(c)),一共发现17个DNB,包括食酸菌属(0.76%~2.06%,Acidovorax)、氢噬菌属(0.23%~ 1.89%,Hydrogenophaga)、Thauera(0.22%~1.77%)、红细菌属(Rhodobacter)、副球菌属(Paracoccus)和红长命菌属(Rubrivivax)等,其中Acidovorax占比最大,其次为Hydrogenophaga和Thauera,Thauera具有反硝化功能,活性污泥体系中Fe3O4 NPs浓度的上升会引起其相对丰度的上升[32]。胁迫阶段,Control中DNB的相对丰度要高于NPs而低于CCZ_L和CCZ_H,可能是3-CCZ与Fe3O4 NPs存在拮抗作用导致复合污染体系中DNB相对丰度的上升,Acidovorax与Hydrogenophaga是其中的优势菌属。低温阶段中各体系Acidovorax相对丰度下降而Hydrogenophaga和Thauera相对丰度则上升,复合污染体系尤其是CCZ_L_2更为明显,Hydrogenophaga和Thauera成为CCZ_L_2中的优势菌属。总体而言,相较于AOB与NOB,DNB受到3-CCZ与Fe3O4 NPs复合污染影响更明显,一些DNB如Hydrogenophaga和Thauera得到了富集。
检测到与硝化作用有关的基因amoA、amoB、amoC和hao(图 4(d)),胁迫阶段中NPs和CCZ_L中这些基因相对丰度皆低于Control,而CCZ_H中相对丰度则高于对照,说明不同浓度3-CCZ与Fe3O4 NPs的复合会对氨氧化基因产生不同影响,高浓度3-CCZ复合污染会促进体系NH4+-N的氧化。而经过低温阶段,除Control_2外所有体系中硝化作用相关基因相对丰度均下降,表明低温下在污染物添加体系中的NH4+-N的氧化较少发生,NH4+-N的去除可能主要有助于其他特定的代谢途径。
将反硝化作用的关键基因分为NO3--N还原基因(narGHI和napAB)和NO2--N还原基因(nirKS、norBC和nosZ)(图 4(d))。在胁迫阶段,添加污染物使得narGHI相对丰度有所减少而napAB则无较大变化,低温阶段NO3--N还原基因相对丰度在各体系内均提高,其中CCZ_L_2中最为明显,而NPs_2和CCZ_H_2中napAB相对丰度均低于对照组。NO2--N还原基因中nosZ的相对丰度在污染物添加后下降并且在低温阶段进一步下降,nirKS与norBC在常温下并未受污染物的影响,但在低温阶段出现了与napAB相似的情况,低温可以使得某些反硝化作用基因的相关丰度增加,低浓度3-CCZ与Fe3O4 NPs的复合污染胁迫下这一情况更明显,可能是低浓度3-CCZ与Fe3O4 NPs之间存在拮抗作用。同时,这也表明在反硝化过程中,具有相似功能的功能基因对复合污染的反应可能不同,Gao等[33]发现norBC和nosZ对阿奇霉素和群体感应抑制剂复合污染胁迫的反应与nirSK相反。总体来说,硝化作用相关功能基因对不同浓度3- CCZ复合污染的反应不同,复合污染胁迫对反硝化功能基因的影响在低温阶段更大,大部分反硝化基因的相对丰度有所上升。
3 结论
1)3-CCZ与Fe3O4 NPs复合污染抑制了活性污泥微生物群落丰富度,但复合污染对群落多样性并没有明显影响。复合污染的添加和低温都会改变微生物群落结构,并且低温阶段中试验组微生物群落结构与对照组差别更大。
2)不同类群微生物对3-CCZ与Fe3O4 NPs复合污染的响应不同,稀有属对污染物的添加更敏感,丰富属中的Saccharimonadales和TM7a一直是优势菌属。CRT与CAT分别是胁迫阶段和低温阶段下微生物交互网络中的核心分类单元,同时稀有属与丰富属之间也存在着复杂的关联。
3)功能微生物中AOB与NOB受复合污染影响较小,DNB的相对丰度则有较大变化尤其在低温阶段,低温下低浓度3-CCZ与Fe3O4 NPs复合污染促进了氢噬菌(Hydrogenophaga)和陶厄氏菌(Thauera)的生长。高浓度3-CCZ与Fe3O4 NPs的复合会引起氨氧化基因的富集,反硝化功能基因nosZ、nirKS和norBC对复合污染产生了不同的反应,nosZ的相对丰度在污染物添加之后下降,后者相对丰度则上升。
综上,常温与低温条件下3-CCZ与Fe3O4 NPs短期复合污染对活性污泥系统丰富-稀有菌属、脱氮功能菌及相关基因的影响存在差异。仍需进一步系统探究不同浓度水平下复合污染长期胁迫下活性污泥系统效能、群落演替、污染物降解机制及耐药菌鉴定等,以期为污水处理系统卤代咔唑与纳米材料复合污染的风险评价及控制提供更为完善的理论依据。







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}