


CO2是一种主要的温室气体,对地球气候系统产生显著影响。开发具有成本效益且可持续的CO2减排方法对于研究国际环境政策至关重要。在现有技术中,微生物固碳以其大规模利用CO2生产化合物、低能耗、无污染的能力脱颖而出[1]。目前大量研究集中在光能自养生物如大型藻类、微藻、蓝藻和紫色细菌上,然而,由于光能自养生物对光的依赖,在生物反应器中扩大其的固碳规模面临重重挑战,包括成本增加以及大型反应器的设计,光穿透率不易控制、热力学效率低和太阳能的间歇性等技术问题。
而吸收CO2的能力并非光合生物所独有,许多化能自养细菌也具备这种能力。它们通过氧化S、S2−、H2S、NH3、NO2−、H2、Fe2+等还原性无机物的过程中获得能量来同化CO2为有机物,进一步合成细胞骨架[2]。化能自养细菌在固碳和碳循环中的重要作用主要归因于以下3个因素:(1) 这些细菌群落通过改变其群落结构、功能范围和生物量水平来响应CO2浓度的变化,使其在极端环境中也能实现固碳[3];(2) 化能自养细菌能够采用专性或兼性营养策略以适应环境中有机物的存在或波动,这种适应性支持它们向生物反应器和工业规模生物过程的潜在转移[4];(3) 化能自养细菌能够将温室气体CO2转化为有价值的化学产品,如生物燃料、生物表面活性剂、胞外多糖和生物聚合物,这种能力不仅显著减少碳排放,还提高了CO2的经济价值[1]。
关于自然生态系统中化能自养细菌固碳的研究激发了其在大规模工业生产中应用的兴趣。已有研究关注在生物反应器中培养和高密度富集化能自养细菌。然而,控制系统与自然生态系统在细菌的生长环境和目标结果方面存在显著差异(表1)。在控制系统中培养和富集化能自养细菌的主要挑战是如何在小体积内实现高细胞密度。要在人工条件下维持固碳的稳定性和效率,必须考虑所有与自然环境不同的过程参数对化能自养细菌高密度培养的影响。在海水等自然生态系统中,微生物群落通过显著的稀释和碳泵效应,缓解了自养细菌生长过程中自我产生的有机代谢产物的积累和环境条件的恶化。然而,在体积有限的生物反应器中复制和放大这些过程是一个重大挑战。
表1 化能自养细菌在自然生态系统与生物反应器中固碳的差异 |
| 区别 | 自然生态系统 | 生物反应器 | |
|---|---|---|---|
| 目标 | 维持生态系统中一定量的有机碳输入 | 扩大细胞培养的规模,提高细胞密度,同时保证系统稳定运行 | |
| 反应体积 | 物质连续流动的广袤的开放系统 | 物质流动不连续且反应体积有限的封闭系统 | |
| 功能微生物 | 微生物群落由隶属于3个生态位的特定微生物组成,包括初级生产、初级降解和最终氧化 | 单一化能自养细菌或人工构建的微生物群落 | |
| 环境条件 | 大自然的缓冲稀释能力可使胞外代谢物浓度维持在较低的水平 | 细菌分泌的有机代谢物在系统中积累,其浓度的逐渐上升使得细菌的生存环境不断恶化 |
尽管研究化能自养细菌在生态系统中固碳取得了进展,但其在人工生物反应器中的固碳潜力在很大程度上被忽视。本文首先介绍了在生物反应器中高密度培养化能自养细菌面临的种种挑战,在此基础上详细讨论了可能的应对对策,最后探讨了在受控环境中高密度培养的化能自养细菌的潜在应用,为未来发展可持续生物生产和负碳微生物经济提供参考(图1)。
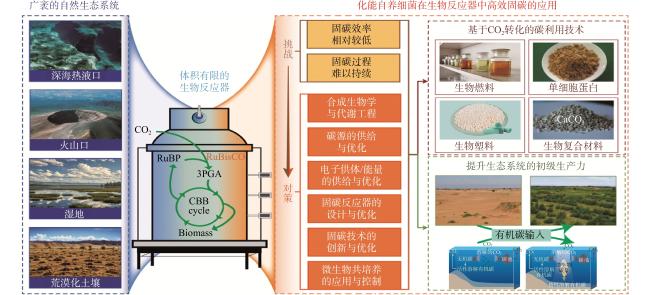
1 化能自养细菌在生物反应器中持续高效固碳所面临的挑战
化能自养细菌独特的碳代谢功能使其在生物反应器中固碳并将CO2转化为高附加值产物成为可能。但影响自养细菌可培养性的因素众多,自然生境下自养细菌细胞的生理状态、营养需求与生态系统中其他生物和非生物因子的相互作用等,均制约着自养细菌细胞是否能够在人为控制条件下生长和繁殖。由于生物反应器体积的有限,一些在自然生境中影响较小的因素(如胞外有机代谢产物的浓度)将在反应器中被放大,从而影响化能自养细菌固碳过程的高效性和可持续性。因此,为了在人工条件下保持其固碳的稳定和高效,必须考虑到不同于自然环境的所有工艺参数对化能自养细菌高密度培养的影响。
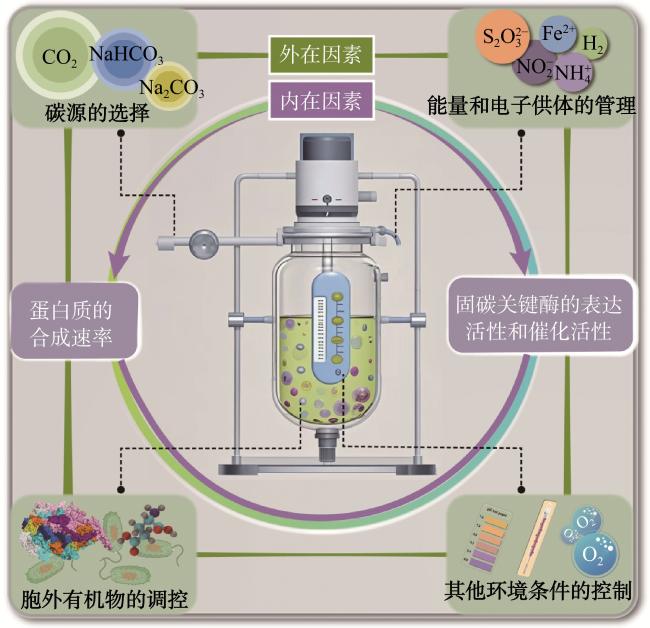
一般而言,所有细菌的生长速率取决于其内在本身的遗传特性及外在的环境条件(图2)。就内部因素而言,化能自养细菌通过氧化还原性物质获得能量和同化CO2获得有机碳为合成细胞骨架提供能量和底物,所以其生长和固碳效率由蛋白质的合成速率和CO2同化途径关键酶的表达活性和催化活性等因素所决定。以卡尔文循环(Calvin−Benson−Bassham cycle,CBB)为例,固碳关键酶RuBisCO催化羧化反应将CO2转化为有机碳化合物,被认为是CO2同化的限速步骤。虽然RuBisCO的I、II和III型参与固碳,但它们同化CO2的速率相对较慢。且当O2存在于反应体系中时,RuBisCO难以区分O2和CO2,即氧可以与CO2竞争相同的结合位点进而抑制化能自养细菌的碳代谢[5]。
由于化能自养细菌通常不能利用有机物,有机物的存在不仅抑制化能自养细菌生长,还会对其固碳过程产生负面影响。有研究证明,某些有机化合物可制约CBB循环中关键酶RuBisCO的合成[13]。尽管某些兼性自养细菌可以在一定程度上同化特定有机化合物,但这种同化能力有限,并不能显著改善其固定CO2的效率。值得注意的是,在生物反应器中易于控制外源有机物,但化能自养细菌仍能在其细胞内合成并释放胞外游离有机物(extracellular free organic carbon,EFOC)。这些溶解有机碳来自细胞内源呼吸过程中内含物的分解和细胞衰亡解体所释放的大分子有机物以及CO2同化过程中的产物,且研究发现这些代谢产物对细胞生长没有有益帮助[14]。尤其在生物反应器中培养化能自养细菌时,要求的细菌密度远高于自然环境,因此,细菌在CO2同化过程中自身产生和积累的EFOC浓度必然较高,若未能及时分离反应器中的EFOC,其积累将显著抑制化能自养细菌的固碳[15]。因此,在人工系统中监控和管理有机物的浓度尤其是EFOC的积累,成为实现高细胞密度和高固碳效率的关键路径和瓶颈。
综上,在生物反应器中通过化能自养细菌实现高效稳定的固碳需要解决多重挑战,不仅需要对细菌内部的代谢过程进行优化,也需要强调在工业应用中外部反应条件的调控,主要包括碳源的选择、能量和电子供体的管理、有机物的影响以及整体环境条件的控制。这些挑战需要通过生物技术手段和工程策略共同应对,以实现工业化规模的化能自养细菌高效固碳。
2 化能自养细菌在生物反应器中持续高效固碳的调控策略
2.1 合成生物学与代谢工程
合成生物学与代谢工程是提高化能自养细菌固碳效率的主要手段之一,目前基于合成生物学与代谢工程的调控主要集中在3个方面(图3):(1) 对天然固碳途径的人工改造和优化。Liang等[16]开发了一种基于活性筛选系统,以实现RuBisCO酶的定向进化,从而提高其羧化活性。此外,优化RuBisCO的CO2/O2选择性也是人工改造的另一策略,通过组装来自不同来源的RuBisCO大、小亚基,可以获得具有高CO2/O2选择性的杂合RuBisCO酶[17]。(2) 利用天然固碳途径中的羧化酶或还原酶设计新的人工固碳途径。例如,Bar−Even等[18]以固碳效率最高的磷酸烯醇式丙酮酸羧化酶为基础,计算获得的合成固碳途径的固碳效率比CBB循环高2~3倍。(3) 寻找新的高效固碳酶以构建人工固碳途径。研究人员利用新酶设计创建了全新的生物固碳途径,包括以CO2为底物合成乙醛酸的巴豆酰辅酶 A/乙基丙二酰辅酶A/羟基丁酰辅酶A(crotonyl−coenzyme A(CoA)/ethylmalonyl−CoA/hydroxybutyryl−CoA,CETCH)循环[19]和不依赖ATP生成乙酰辅酶A的合成乙酰辅酶A(Synthetic Acetyl−CoA,SACA)固碳途径[20]。值得注意的是,凭借单一的优化方案提高固碳效率的效果是有限的,未来仍需合理设计多途径优化以提高工业应用中微生物固碳的效率。
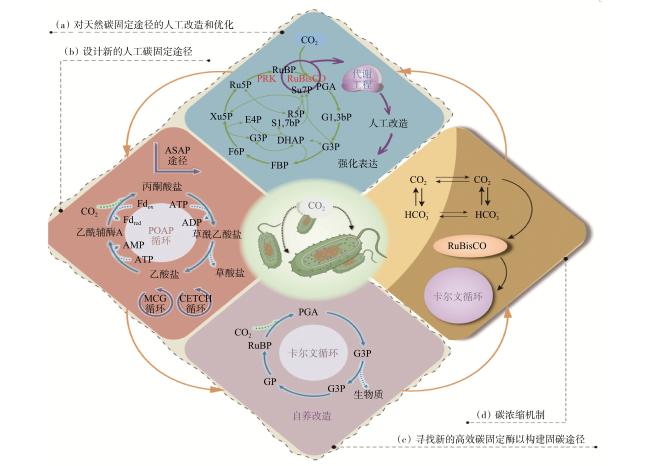
此外,微生物内部的CO2浓缩机制(CO2−concentrating mechanism,CCM)是提高固碳酶催化速率的常用策略[21],目的在于克服CO2在水中的低扩散速率和改善细胞内部固碳酶RuBisCO周围的CO2浓度,从而提高固碳效率。目前,在CO2的生物固定中,CCM系统的在化能自养细菌中改造和应用主要包括2个方面:(1) 无机碳传输系统的异源重建。提高酶性能的常见策略是增加底物浓度以提高催化速率,尤其是CO2浓度。Xiao等[22]通过过表达Synechococcus sp. PCC 7002细菌中的碳酸氢盐转运蛋白基因bicA,发现与对照菌株相比固碳效率提高了59.3%。该研究预示了在固碳微生物中重建无机碳传输系统在提高固碳效率方面的有效性。(2) 羧基小体的工程改造。羧基体的独特结构和半渗透性使其在催化增强、酶保护和分子递送方面具备应用潜力,将酶掺入蛋白质壳中,以进行区室化和提高催化活性[23]。构建含有固碳酶、碳酸酐酶或其他酶的功能性人工羧基体以取代原始RuBisCO,可能在未来进一步提高化能自养细菌的固碳效率。
2.2 碳源的供给与优化
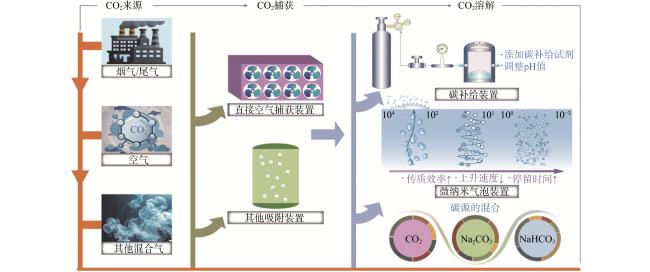
首先,近年来捕获和浓缩CO2的方法引起了广泛关注。直接空气捕获(direct air capture,DAC)技术可直接从环境中捕获CO2,再通过向捕获介质中引入能量可释放浓缩的CO2[26]。Sillman等[27]评估了使用DAC生产微生物蛋白的潜力,发现利用空气捕获和可再生能源进行微生物蛋白生产所需的土地和水比大豆生产更少。然而,这种方法的技术和经济可行性需要仔细评估,尤其是考虑到大规模应用DAC所需的额外能量消耗和成本。此外,如果在废气进入固碳反应器之前保留CO2并去除抑制细菌生长的杂质,可以低成本地增强细菌的固碳。一些化能自养细菌可以从烟气中的硫化物和氧化铁中获取能量,同时使用烟气中的CO2作为碳源[28]。然而,该方法仍面临包括提高气液传质效率和去除烟气中如SO2、NOx、CH3OH和粉尘等抑制细菌生长的杂质技术在内的重重挑战[29−30]。
其次,改善碳源溶解度的方法包括开发更高效的生物反应器、设计碳补给装置、调整pH值和添加碳补给试剂。Bao等[31]设计了一种原位碳补给系统,在开放式跑道池反应器中将平均CO2吸收率和CO2利用率分别提高到86.2%和79.2%。Rosa等[32]发现,与NaOH相比,单乙醇胺可以将藻类液体中的总无机碳浓度提高一倍,并使生物质蛋白浓度增加17%。微纳米气泡技术是一种取得显著进展的创新方法。该技术通过将气体驱入水中形成微纳米气泡,在水中气泡的收缩和溶解过程中实现过饱和状态[33]。Guo等[34]利用该技术增强海洋微生物Sinomicrobium oceani WH−15的生长和代谢,观察到微纳米气泡通过增加传质、降低氧化还原电位、增加无机碳源的可用性和减少胞外代谢物等机制促进了微生物的CO2固定。
2.3 电子供体/能量的供给与优化
充足的能量供应是提高化能自养细菌固碳效率的关键瓶颈。电子供体如H2和无机盐向电子受体如氧气的电子转移主要依赖于微生物体内的还原能量供应。选择合适的电子供体对于微生物固碳至关重要,电子转移速率是限制其效率的关键因素。因此,有必要基于机制创新来优化电子供应方法(图5)。
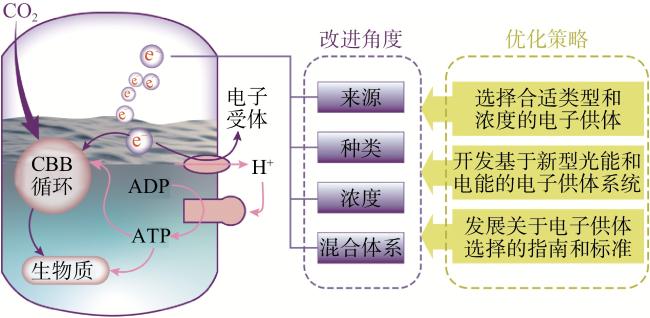
为了应对固碳过程中代谢还原力和能量供应不足的问题,可开发利用光能和电能的多种电子供体系统。Sakimoto等[40]和Zhang等[41]设计了一个结合太阳能与无机材料用于化学合成的混合系统。在这个系统中,半导体纳米颗粒被沉积在表面,作为电子收集器,为化能自养的温热酵母供应电子以固定CO2。此外,电子活性微生物可以通过胞外电子传递链或导电蛋白直接从阴极接受电子,提供固碳所需的还原力。例如,产乙酸菌Clostridium ljungdahlii DSM 13528可以直接接受阴极电子,使乙酸产生量增加3倍[42]。材料科学和系统生物学的进步不断改进电子介质的性能、光能材料的转换效率以及生物系统的兼容性。结合元件(电子介质和电子传递器)和关键步骤(还原力保持和能量再生)的研究仍需进一步细化。
2.4 反应系统优化以消除积累的有机代谢产物的反馈抑制
就化能自养菌的固碳过程而言,将微生物反应过程与EFOC分离有利于化能自养细菌在有限体积的生物反应器中的持续固碳。研究人员不断探索新的生物技术方法来管理代谢物积累,这在工业应用和生物技术中具有重要的实际意义(图6)。
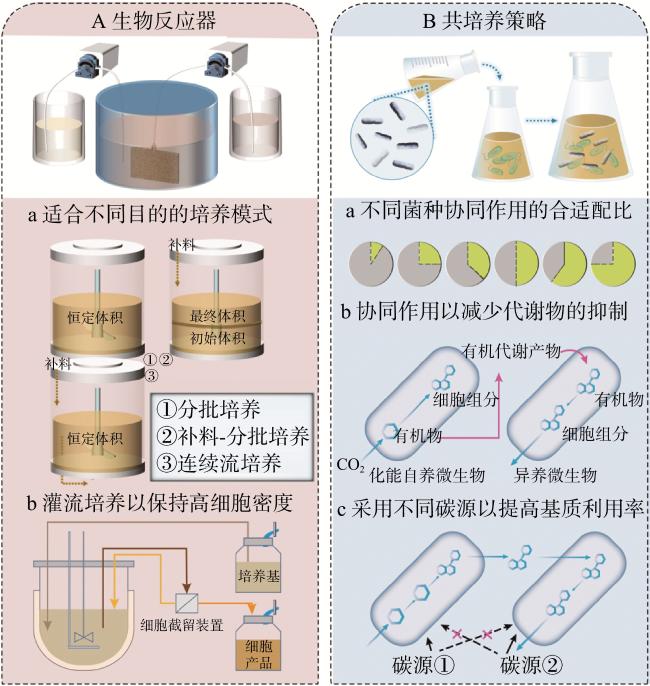
分批培养是指在开始时添加培养基和细菌,在结束时收获产品。然而,由于细菌分泌的有机代谢产物在体积有限的反应器中不断积累及其对细菌产生反馈抑制[13],分批培养无法维持化能自养细菌的持续生长和固碳。相较而言,连续培养过程提供了更大的操作灵活性和更高的生产率,通过培养基的不断流入和输出使胞外积累的有机代谢产物浓度降低。然而,工业生产必须考虑培养基更换速度远高于细菌生长周期带来的细菌流失,因此考虑将连续流培养过程与截留细菌相结合。Zhang等[43]设计了一种连续流膜生物反应器使其在去除胞外有机代谢产物的同时通过膜组件截留随培养基流出的细菌,结果表明,膜反应器中累积的表观固碳效率是对照反应器的3.24倍。更进一步地,由于生命周期评估分析强调环境影响是未来集约化生产的一个重要问题,因此,在培养基流出过程中,有必要采用一些策略来回收培养基并回收潜在的副产品,以最大限度地减少对环境的影响。Wan等[44]开发了一种用于自动灌注细胞培养的反应器,该反应器可以有效分析营养物质、代谢产物和细胞浓度。该反应器系统在持续刷新生物反应器内培养基的同时去除使用过的培养基和细胞废物。通过提供降低培养基成本或优化培养基的策略来解决培养基成本的瓶颈问题,包括培养基回收新技术的开发、细胞代谢产生的废物的评估以及生长因子特性的改善等。
除了膜过滤之外,其他分离剂或技术也常用于生物反应与代谢物分离的结合过程中。可以采用如透析、吸附或特定树脂等技术来去除培养基中的抑制副产物。例如,去除发酵过程中产生的有机酸可以缓解对细菌培养的压力,允许持续生长[45]。Fuchs等[46]发现,通过透析发酵从细胞悬浮液中去除抑制代谢物可以延长指数生长期并产生更高产量,细胞密度超过190 g/L。此外,Zhao等[47]证明了使用低释放生物炭吸附化能自养细菌硫杆菌的胞外代谢物促进微生物生长190%,增加CO2固定29%。这些技术在去除胞外代谢物和增强细菌生长方面提供了显著的益处。技术的选择取决于具体的过程要求,包括成本、可扩展性和目标代谢物的性质。
除了物理分离胞外代谢物,共培养是一种有效的策略来消除此类反馈抑制。Kerckhof等[48]从9株甲烷氧化菌(methane−oxidizing bacteria,MOB)菌株和19株氢氧化细菌(hydrogen−oxidizing bacteria,HOB)菌株中筛选出最优的共培养组合,其蛋白质浓度是单独HOB菌株生产量的3.8倍。Hu等[49]将HOB与异养微生物共培养,获得了生物量增加20%,蛋白质增加24%,必需氨基酸生产增加28%。为了模拟环境中的微生物相互作用并促进培养,开发了专门的共培养反应器。例如,Burmølle等[50]创建了一种双层介质,与单层介质相比,产生的菌落多出20%~40%。此外,Acosta等[51]将MOB和HOB的共培养与水电解和沼气发酵相结合,最终提高了17%的蛋白质生产。在这种设置中,MOB氧化CH4产生的CO2被送入HOB。
在自然生态系统中,自养和异养微生物通常形成维持这些生态系统固碳功能的共生群落。然而,在生物反应器中复现这种共生关系仍具挑战性,需要进一步研究以提供科学依据。一些问题仍需进一步探明,例如,固碳是由整个微生物群落驱动的,还是自养和异养微生物共同协作的结果,抑或自养微生物起主导作用。此外,不同自养固碳途径对微生物群落综合作用的响应尚不清楚,还需要进一步研究以阐明共培养系统中化能自养菌和异养菌的碳同化机制,包括对固碳过程功能基因的分析——特别是基因组、转录组、蛋白组和代谢组分析,以全面探索潜在的新碳同化机制。
3 生物反应器中高密度培养的化能自养固碳细菌的应用前景
3.1 提升低有机碳输入生态系统的初级生产力
深海化能自养固碳(dark carbon fixation, DCF)因其作为海洋食物网(包括水柱和沉积物)有机物的重要来源而备受关注[55]。传统上,人们认为光能自养生物固碳是海洋生产力的主要方式,其在真光层产生的颗粒有机碳满足了深海无光区的能量需求[56]。然而,近年来发现海洋生物对能量的需求超过了颗粒有机碳的能量输入,并且随着深度的增加,海洋能量收支的不平衡变得更加明显[57]。已有研究表明,海洋学家估计全球海洋化能自养碳固定总量约为0.77 PgC/a,这表明,化能自养生物仍然可以与光自养生物竞争海洋透光层中溶解的无机碳,并进行有效的趋化自养碳固定[58]。未来,随着海洋持续升温,海洋生产力与颗粒有机物输出通量可能减少。因此,向深海生物圈人工输入化能自养细菌来支持DCF可以补充透光层的能量输入的不足,增加细菌有机质对沉降颗粒有机物输出通量的贡献和与之相关的难降解溶解有机质生产和输出。
土壤是陆地生态系统中最大的碳库,也是地表生态系统中最活跃的碳库之一。研究表明,化能自养菌能够利用CO2作为碳源合成自身营养物质,对土壤固碳有重要贡献[59]。标记实验直接追踪固定的CO2到土壤有机碳(soil organic carbon,SOCs),表明沙漠土壤吸收的大气CO2中近4%被合并到SOCs中,这支持了微生物CO2同化在沙漠土壤中SOCs形成的作用[60]。此外,土壤中的自养微生物在低养分条件下往往表现出更高的净固碳率[61]。适应贫营养环境、干旱和极端温度的化能自养细菌通过固碳合成有机物,从而缓解了主要异养微生物的环境胁迫[62]。通过研究微生物同化碳在土壤碳库中的转化过程和稳定机制,可将碳同化微生物在土壤CO2同化过程中的贡献定量化,在此基础上,可以采取一系列人工手段提升微生物介导的土壤碳过程,采取合理有效的措施提升土壤有机碳含量和土壤生产力功能,为农业生产增收增益。
综上,化能自养细菌的固碳潜力对于稳定海洋内部、沙漠土壤和其他低有机碳输入生态系统的碳循环至关重要。理论上,将固定的无机碳转化为有机生物聚合物可以增强整体固碳。通过人工引入化能自养细菌以增强微生物有机物输入,低有机碳输入生态系统的初级生产力可以有效恢复。然而,化能自养菌在生态系统中固碳的应用大多是基于基因组预测或实验室规模实验推测的,尚未通过现场实验验证。化能自养菌对异养过程、初级社区的微生物动态以及生态系统中微生物食物网的形成和演替的影响尚不清楚。未来的研究应结合现代技术,如异源表达、蛋白质组学和代谢组学,以确定化能自养菌在现场碳转换的活性和速率。同时,在实验室中分离和构建合适的化能自养菌并测试其生态潜力至关重要。
3.2 基于CO2生物转化的高价值产品生产
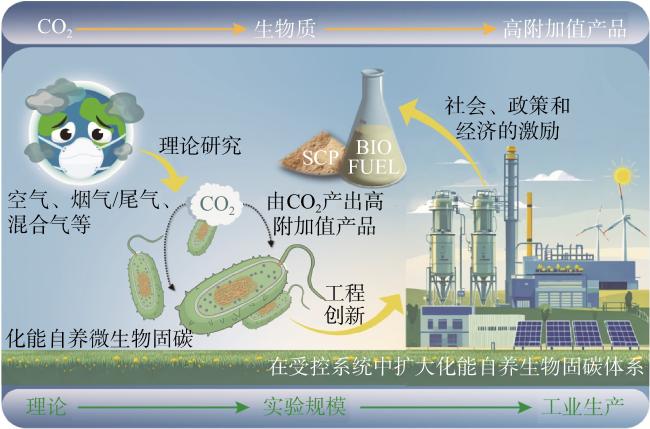
除了生物燃料,单细胞蛋白(single cell protein, SCP)因其高蛋白含量和营养价值越来越受到关注。世界经济论坛第十五届新领军者年会(夏季达沃斯论坛)发布年度《十大新兴技术报告》,其中就包括碳捕获微生物和牲畜饲料蛋白替代技术[69]。目前,大多数SCP是由异养细菌生产的,这些细菌以碳水化合物、脂类或如CH4和CH3OH等底物生长,在此过程中会导致CO2的产生。相较而言,化能自养细菌的CO2同化能力使其成为真正可持续SCP生产的最佳候选者,通过化能自养细菌将CO2转化为SCP可以为农业提供可持续的解决方案。目前,Clostridium ethanolis生产的微生物蛋白已用于动物饲料,并有望成为新型替代食品蛋白,但需要在生产过程中加入核酸去除步骤[70]。氢氧化细菌还可以通过在生物反应器中利用水电解产生的H2作为能源生产单细胞蛋白[71−72]。然而,这些细菌的生长和培养需要持续的氢气供应,存在风险且生产成本高。何环等[73]的研究指出,在单质硫中生长的细胞及分离得到的胞外蛋白中富含巯基的蛋白含量明显增加,这表明硫氧化细菌可能是一种潜在的高质量蛋白的生产者。因此,通过化能自养细菌高效转化CO2为单细胞蛋白是值得关注的一个新的应用方向,进一步研究可从高效菌株的选择、自养微生物蛋白生产的影响因素和机制,以及调控和优化转化过程等关键领域展开。
综上,收获的化能自养细菌生物量可以用于生产高附加值产品,代替现有的商业产品,从而减少对合成材料的依赖。然而,在工业规模上使用这些生物进行碳利用仍然面临一系列挑战,如生物量产量低、高生物反应器维护成本以及从生物量中收获和提取材料的费用等。此外,消费者的接受度显著影响生产过程的经济可行性,对于生物产品的有效商业化至关重要。
4 结论与展望
化能自养细菌通过将无机碳转化为有机碳从而参与全球碳循环过程。将其固碳特性于工业规模加以应用具有可观的生态和经济效益,但这就不得不考虑在生物反应器中高密度培养细菌面临的一系列挑战。一个长期目标是根据合成生物学原则驯化新的宿主进行代谢工程,以创建完全高效的细胞工厂。实验实施还需要测试不同系统的控制方案以减少环境影响并提高能源资源效率,这包括优化营养来源和削弱代谢产物的反馈抑制。富集化能自养细菌不仅可以利用温室气体作为碳源,收获的菌体还可还至低有机物输入的生态系统以提高其初级生产力,也可用于炼制高价值产品以抵消固碳成本并产生经济效益。
期望未来整合得到验证的技术和方法,克服在生物反应器中培养化能自养细菌的瓶颈,开发评估碳转化产品经济价值的标准,并进行全面的固碳核算和生命周期评估。同时,生物技术工程必须解决生产力、安全性和资本强度等问题,通过新的生产策略开发用于化能自养细菌的CO2捕获和转化的新系统。此外,应补充选择绿色资源再生技术,并构建适合高附加值产品的应用场景,以支持高效碳回收和增值利用的低碳范式的发展。

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}